

Essa é uma revisão anterior do documento!
Tabela de conteúdos
Ensaios 2022
Resiliência ecológica em agroecossistemas
Julie Christine Scaloppi
O conceito de resiliência, utilizado em diversas áreas do conhecimento, pode ser empregado na ecologia a partir de diferentes abordagens (Holling 1996, Gunderson 2000, Folke 2006). Quando se considera que sistemas dinâmicos podem exibir estados estáveis alternativos, resiliência pode ser conceituada como a capacidade do sistema persistir em uma bacia, ou domínio de atração, frente à eventuais pertubações (Holling 1973). O estado em que o sistema se encontra pode ser compreendido como um balanço de processos. Estados estáveis são aqueles onde o balanço de processos tende a manter o sistema em um equilíbrio dinâmico, ou seja, os processos que influenciam o sistema resultam em forças que contribuem para sua manutenção, formando um domínio ou bacia de atração (Scheffer 2009). Diante de determinada intensidade ou frequência de perturbações, as forças atratoras de um estado estável podem não ser suficientes para preservar o sistema naquela bacia de atração. Quando o sistema atinge o limite de uma bacia de atração ele chega à uma bifurcação, um estado instável a partir do qual uma pequena perturbação adicional pode provocar uma transição crítica, onde o sistema entra em uma bacia de atração de um estado estável alternativo (May 1977, Scheffer et al. 2001).
A resiliência no contexto dos sistemas com múltiplas bacias de atração, denominada resiliência ecológica, é uma propriedade que representa a magnitude de distúrbio que o sistema consegue absorver sem mudar de estado (Holling 1973). O conceito é relevante para a caracterização dos estados de sistemas ecológicos. A relação entre a diversidade biológica e a resiliência é um tema de interesse da teoria ecológica (Gunderson 2000). Maiores diversidades de organismos possibilitam maiores diversidades e redundâncias funcionais nas comunidades ecológicas. Hipóteses que relacionam positivamente a biodiversidade e resiliência ecológica sugerem que essas características reduzem a susceptibilidade das comunidades à perturbações. Assim, a complementaridade no uso de recursos, a diversidade de desempenhos e o fato da perda de espécies ser menos nociva ao sistema como um todo, reduzem a fragilidade e aumentam a responsividade do sistema à perturbação (Elmqvist et al. 2003, Downing et al. 2012). Nesse sentido, a caracterização da diversidade e redundância funcional das comunidades poderiam indicar se o sistema se encontra em um estado estável ou próximo à uma transição crítica (Roberts et al. 2019). O conceito de resiliência ecológica também considera a capacidade do sistema de absorver distúrbios e se reorganizar, mantendo sua função e estrutura. Além da importância teórica, a resiliência ecológica contribui para a compreensão dos mecanismos que mantêm estados estáveis e para a identificação de sinais de aproximação das transições críticas dos ecossistemas, fundamental para a conservação e manejo de sistemas naturais (Scheffer et al. 2001, Dakos et al. 2015).
Sistemas ecológicos caracterizados por estados estáveis alternativos, induzidos ou não por atividades antrópicas, têm sido observados em diversos ecossistemas aquáticos (Blindow et al. 1993, Knowlton 2004, Roelke et al. 2007, Carpenter et al. 2011) e terrestres (Dublin et al. 1990, Walker et al. 1997, Warman and Moles 2009, Eby et al. 2017, Katembo et al. 2020). No caso do lago Takvatn, localizado na Noruega, analisado por Persson et al. (2007), a mudança de estado foi atribuída à pressão das atividades de pesca. Um manejo contraintuitivo, baseado nas formas pelas quais a predação seletiva em relação ao tamanho de presas e as características das espécies de peixes do lago poderiam causar estados alternativos de equilíbrio, auxiliou na reversão e manutenção do estado onde a espécie comercial de interesse tem sua população aumentada. O estudo também sugere que o acompanhamento da população das presas poderia indicar riscos de colapso desse sistema. O estudo de Persson et al. (2007) e sua sugestão de manejo, contrasta com a abordagem de “rendimento máximo sustentável” aplicado por indústrias de pesca ao redor do mundo. O indicador de limite de exploração utilizado por essa abordagem está relacionado somente com a população de interesse comercial, o que implica em enorme chances de colapso das comunidades aquáticas onde se inserem essas populações de interesse (Legovic et al. 2010). É importante destacar que a mudança e reversão de estados estáveis de um sistema pode ser caracterizada por condições críticas distintas, ou seja, a mudança de estado passa por um estado instável distinto do percorrido durante a reversão dessa mudança. Nesse caso, o sistema é caracterizado pela histerese. Se o sistema apresenta histerese e, por exemplo, o novo estado é indesejado, a reversão para o estado anterior pode exigir muito mais do que a reversão das perturbações ou a restauração das condições anteriores (Beisner et al. 2003, Scheffer 2009). Um exemplo são as transições provocadas por pressões de pastejo que levam ecossistemas áridos à estados desertificados (Kéfi et al. 2007).
As atividades antrópicas interferem nos processos que controlam os fluxos de energia, nutrientes e matéria orgânica nos sistemas ecológicos, as funções ecossistêmicas. Muitas vezes, essas alterações têm como consequência a redução na biodiversidade (Cardinale et al. 2012). Aproximadamente um quarto da superfície terrestre foi transformada em sistemas cultivados para atender as crescentes demandas por alimento, madeira, fibras e combustíveis. A distribuição das espécies tem se tornado mais homogênea e a diversidade genética global está sendo reduzida, uma das razões é a padronização das variedades cultivadas. Além disso, a demanda por água, fertilizantes e combustíveis fósseis das atividades produtivas têm alterado os processos e funcionamentos dos ciclos biogeoquímicos (MEA 2005). A teoria ecológica associada a dados empíricos tem mostrado que a redução de diversidade diminui a estabilidade temporal das funções ecossistêmicas, como a produção de biomassa, a eficiência da captura de recursos, a decomposição e a ciclagem de nutrientes nos sistemas ecológicos (Cardinale et al. 2012). Uma das estratégias propostas para atenuar esses impactos consiste na diversificação dos agroecossistemas. A transição de agroecossistemas simplificados para agroecossistemas mais biodiversos têm sido apontada como prioridade no contexto de perda de biodiversidade mundial, especialmente devido ao potencial de otimizar as áreas produtivas, reduzir a conversão de novas áreas para produção agrícola e aumentar a biodiversidade nas áreas já convertidas (Malézieux et al. 2009, Vandermeer 2011, Sachs et al. 2019).
A aplicação do conceito de resiliência ecológica em agroecossistemas deve considerar os objetivos e limitações inerentes a esses sistemas, que são distintos dos sistemas naturais. Para isso, Peterson et al. (2018) recomendam que a análise da resiliência ecológica em agroecossistemas utilize uma estrutura onde as funções produtivas, os mecanismos regulatórios e as escalas sejam explicitadas. Os mecanismos regulatórios que aumentam resiliência de determinado estado podem ser externos ou internos ao agroecossistema. Sistemas regulados com mecanismos externos geralmente possuem produtividade centrada em uma única escala e intervalo de tempo e são altamente dependentes de insumos externos. Por sua vez, agroecossistemas regulados através de mecanismos internos se baseiam na diversidade genética e de espécies, estruturas tróficas complexas e acumulação de matéria orgânica (Peterson et al. 2018). Nesse sentido, o aumento da diversidade pode funcionar como um mecanismo endógeno de agroecossistema, que aumenta a resiliência de um estado estável desejado e também permite a manutenção das funções ecossistêmicas frente a estresses e mudanças ambientais (Oliver et al. 2015, Martin et al. 2019).
As interações entre espécies de plantas em agroecossistemas biodiversos são o foco da minha pesquisa. O conceito de resiliência ecológica contribui para uma perspectiva na qual o aumento da biodiversidade pode favorecer a manutenção de estados desejados em agroecossistemas. As comunidades de plantas inseridas e cultivadas nos agroecossistemas são consequência da manipulação e manejo antrópico, porém, influenciadas pelos mesmos mecanismos que conduzem a estrutura e dinâmica nos ambientes naturais (Vandermeer 2011). As vantagens associadas ao aumento da biodiversidade em áreas agrícolas dependem da escolha e combinação de organismos e de traços funcionais. A escolha, que deve considerar as condições ambientais locais, busca promover a complementaridade no uso de recursos, com ênfase em processos de facilitação entre organismos (Cardinale et al. 2002, Liu et al. 2018). Entre as vantagens estão o melhor aproveitamento dos recursos disponíveis e, potencialmente, melhorias no microclima e favorecimento da polinização e controle de pragas e doenças, resultantes de interações diretas e indiretas entre espécies (Brooker et al. 2015). O estudo das interações entre plantas em agroecossistemas é uma oportunidade de desenvolvimento de sistemas mais resilientes, produtivos e que realizem a manutenção das funções ecossistêmicas (Brooker et al. 2021).
Referências bibliográficas
Beisner, B. E., D. T. Haydon, and K. Cuddington. 2003. Alternative stable states in ecology. Frontiers in Ecology and the Environment 1:376–382.
Blindow, I., G. Anderson, A. Hargeby, and S. Johansson. 1993. Long‐term pattern of alternative stable states in two shallow eutrophic lakes. Freshwater Biology 30:159–167.
Brooker, R. W., A. E. Bennett, W. F. Cong, T. J. Daniell, T. S. George, P. D. Hallett, C. Hawes, P. P. M. Iannetta, H. G. Jones, A. J. Karley, L. Li, B. M. Mckenzie, R. J. Pakeman, E. Paterson, C. Schöb, J. Shen, G. Squire, C. A. Watson, C. Zhang, F. Zhang, J. Zhang, and P. J. White. 2015. Improving intercropping: A synthesis of research in agronomy, plant physiology and ecology. New Phytologist 206:107–117.
Brooker, R. W., T. S. George, Z. Homulle, A. J. Karley, A. C. Newton, R. J. Pakeman, and C. Schöb. 2021. Facilitation and biodiversity–ecosystem function relationships in crop production systems and their role in sustainable farming. Journal of Ecology 109:2054–2067.
Cardinale, B. J., J. E. Duffy, A. Gonzalez, D. U. Hooper, C. Perrings, P. Venail, A. Narwani, G. M. MacE, D. Tilman, D. A. Wardle, A. P. Kinzig, G. C. Daily, M. Loreau, J. B. Grace, A. Larigauderie, D. S. Srivastava, and S. Naeem. 2012. Biodiversity loss and its impact on humanity. Nature 486:59–67.
Cardinale, B. J., M. A. Palmer, and S. L. Collins. 2002. Species diversity enhances ecosystem functioning through interspecific facilitation. Nature 415:426–429.
Carpenter, S. R., J. J. Cole, M. L. Pace, R. Batt, W. A. Brock, T. Cline, J. Coloso, J. R. Hodgson, J. F. Kitchell, D. A. Seekell, L. Smith, and B. Weidel. 2011. Early warnings of regime shifts: A whole-ecosystem experiment. Science 332:1079–1082.
Dakos, V., S. R. Carpenter, E. H. van Nes, and M. Scheffer. 2015. Resilience indicators: Prospects and limitations for early warnings of regime shifts. Philosophical Transactions of the Royal Society B: Biological Sciences 370:1–10.
Downing, A. S., E. H. van Nes, W. M. Mooij, and M. Scheffer. 2012. The resilience and resistance of an ecosystem to a collapse of diversity. PLoS ONE 7:1–7.
Dublin, H. T., A. R. E. Sinclair, and J. McGlade. 1990. Elephants and fire as causes of multiple stable states in the Serengeti-Mara woodlands. Journal of Animal Ecology 59:1147–1164.
Eby, S., A. Agrawal, S. Majumder, A. P. Dobson, and V. Guttal. 2017. Alternative stable states and spatial indicators of critical slowing down along a spatial gradient in a savanna ecosystem. Global Ecology and Biogeography 26:638–649.
Elmqvist, T., C. Folke, M. Nyström, G. Peterson, J. Bengtsson, B. Walker, and J. Norberg. 2003. Response diversity, ecosystem change, and resilience. Frontiers in Ecology and the Environment 1:488–494.
Folke, C. 2006. Resilience: The emergence of a perspective for social-ecological systems analyses. Global Environmental Change 16:253–267.
Gunderson, L. H. 2000. Ecological Resilience - in theory and application. Annual Review of Ecology and Systematics 31:425–439.
Holling, C. S. 1973. Resilience and stability of ecological systems. Annual Review of Ecology and Systematics 4:1–23.
Holling, C. S. 1996. Engineering Resilience versus Ecological Resilience. Pages 31–44 in P. C. Schulze, editor. Engineering Within Ecological Constraints. Nacional Academy Press, Washington.
Katembo, J. M., M. B. Libalah, F. B. Boyemba, G. Dauby, and N. Barbier. 2020. Multiple stable dominance states in the congo basin forests. Forests 11:1–16.
Kéfi, S., M. Rietkerk, C. L. Alados, Y. Pueyo, V. P. Papanastasis, A. ElAich, and P. C. De Ruiter. 2007. Spatial vegetation patterns and imminent desertification in Mediterranean arid ecosystems. Nature 449:213–217.
Knowlton, N. 2004. Multiple “stable” states and the conservation of marine ecosystems. Progress in Oceanography 60:387–396.
Legovic, T., J. Klanjšček, and S. Geček. 2010. Maximum sustainable yield and species extinction in ecosystems. Ecological Modelling 221:1569–1574.
Liu, C. L. C., O. Kuchma, and K. V. Krutovsky. 2018. Mixed-species versus monocultures in plantation forestry: Development, benefits, ecosystem services and perspectives for the future. Global Ecology and Conservation 15:e00419.
Malézieux, E., Y. Crozat, C. Dupraz, M. Laurans, D. Makowski, H. Ozier-Lafontaine, B. Rapidel, S. de Tourdonnet, and M. Valantin-Morison. 2009. Mixing plant species in cropping systems: concepts, tools and models. A review. Agronomy for Sustainable Development 29:43–62.
Martin, E. A., B. Feit, F. Requier, H. Friberg, and M. Jonsson. 2019. Assessing the resilience of biodiversity-driven functions in agroecosystems under environmental change. Pages 59–123 in D. A. Bohan and A. J. Dumbrell, editors. Advances in Ecological Research. First edition. Academic Press.
May, R. M. 1977. Thresholds and breakpoints in ecosystems with a multiplicity of stable states. Nature 269:471–477.
MEA, M. E. A. 2005. Ecosystems and human well-being: Biodiversity Synthesis. Page (J. Sarukhán and A. Whyte, Eds.). World Resources Institute, Washington.
Oliver, T. H., M. S. Heard, N. J. B. Isaac, D. B. Roy, D. Procter, F. Eigenbrod, R. Freckleton, A. Hector, C. D. L. Orme, O. L. Petchey, V. Proença, D. Raffaelli, K. B. Suttle, G. M. Mace, B. Martín-López, B. A. Woodcock, and J. M. Bullock. 2015. Biodiversity and Resilience of Ecosystem Functions. Trends in Ecology and Evolution 30:673–684.
Peterson, C. A., V. T. Eviner, and A. C. M. Gaudin. 2018. Ways forward for resilience research in agroecosystems. Agricultural Systems 162:19–27.
Persson, L., P. A. Amundsen, A. M. De Roos, A. Klemetsen, R. Knudsen, and R. Primicerio. 2007. Culling prey promotes predator recovery - Alternative states in a whole-lake experiment. Science 316:1743–1746.
Roberts, C. P., D. Twidwell, D. G. Angeler, and C. R. Allen. 2019. How do ecological resilience metrics relate to community stability and collapse? Ecological Indicators 107:105552.
Roelke, D. L., T. Zohary, K. D. Hambright, and J. V. Montoya. 2007. Alternative states in the phytoplankton of Lake Kinneret, Israel (Sea of Galilee). Freshwater Biology 52:399–411.
Sachs, J. D., G. Schmidt-Traub, M. Mazzucato, D. Messner, N. Nakicenovic, and J. Rockström. 2019. Six Transformations to achieve the Sustainable Development Goals. Nature Sustainability 2:805–814.
Scheffer, M. 2009. Alternative Stable States. Pages 11–36 in M. Scheffer, editor. Critical Transitions in Nature and Society. Princeton University Press, Princeton.
Scheffer, M., S. Carpenter, J. A. Foley, C. Folke, and B. Walker. 2001. Catastrophic shifts in ecosystems. Nature 413:591–596.
Vandermeer, J. H. 2011. Competition and Facilitation Among Plants: Intercropping, Weeds, Fire, and the Plow. Pages 63- in J. H. Vandermeer, editor. The Ecology of Agroecosystems. Jones and Bartlett Publishers, Sudbury.
Walker, B. H., J. L. Langridge, and F. McFarlane. 1997. Resilience of an Australian savanna grassland to selective and non-selective perturbations. Austral Ecology 22:125–135.
Warman, L., and A. T. Moles. 2009. Alternative stable states in Australia’s Wet Tropics: A theoretical framework for the field data and a field-case for the theory. Landscape Ecology 24:1–13.
A competição e a partilha de recursos em guildas tróficas de aves sob a ótica dos isótopos estáveis
Ana Beatriz Navarro
O conceito da competição é um dos mais explorados historicamente dentro da Ecologia de Comunidades, levando a existência de uma série de estudos envolvendo modelos tanto de caráter fenomenológico (modelo de Lotka-Volterra) quanto mecanístico (modelo de Tilman). A conceitualização geral do processo de competição em uma comunidade nada mais representa que a interação entre indivíduos (sejam eles da mesma espécie ou de espécies diferentes) que compartilham a necessidade por um mesmo recurso limitado (e.g., alimento, nutriente, habitat). De modo amplo, existem dois mecanismos de competição interespecífica que podem operar em uma comunidade, (1) a competição exploitativa e (2) a competição de interferência (Morin, 2011). A competição exploitativa ocorre quando uma espécie partilha de um mesmo recurso com outra espécie, limitando o crescimento populacional de ambas ao tornar o recurso em questão menos disponível. Já a competição de interferência ocorre quando uma espécie limita o acesso de outra espécie a um recurso comunitário, o que pode envolver territorialidade ou defesas químicas. Assim, independentemente do tipo de competição em progresso em uma comunidade, é fato que tal processo leva a uma interação negativa entre os indivíduos competidores, regulando o tamanho populacional das espécies através de uma redução na taxa de crescimento e, assim, modulando a estruturação da comunidade.
A importância histórica do conceito da competição para a ecologia remonta à década de 1920, quando Alfred J. Lotka (em 1924) e Vito Volterra (em 1926) deduzem de forma independente um modelo matemático que descreve as interações competitivas entre populações de duas espécies e as condições necessárias para a coexistência das mesmas. Ademais, o conceito da competição exerce forte influência na década de 1930, quando Georgy F. Gause, através de experimentos com protozoários do gênero Paramecium, formula o princípio da exclusão competitiva (também conhecido como ‘princípio de Gause’), que demonstra a impossibilidade de coexistência entre espécies com nichos similares (Gause, 1934). Segundo esse princípio, uma espécie X, com melhor habilidade competitiva por determinado recurso limitado, excluirá outra espécie Y, também dependente do recurso, visto que Y é competitivamente inferior. A coexistência entre as espécies só seria bem sucedida caso houvesse uma partilha de recursos, ou seja, que as espécies apresentassem uma diferenciação em seus nichos. Essa possibilidade de coexistência estável entre espécies competidoras pode ser deduzida utilizando-se o modelo de Lotka-Volterra, segundo a seguinte equação: (1) dN1/dt = r1*N1 [(K1-N1-α12*N2) / K1], em que dN1/dt é a taxa de crescimento da população 1 (sendo igual a 0 quando a população está no equilíbrio, ou seja, o número de indivíduos permanece constante ao longo do tempo), r1 é a taxa intrínseca de crescimento natural da população 1, N1 é o tamanho da população 1, K1 é a capacidade de suporte da população 1, α12 é o coeficiente de competição da espécie 2 sobre 1, e N2 é o tamanho da população 2. Sabemos que, se α > 1 a competição interespecífica é mais importante para a regulação da espécie em questão e se α < 1 a competição intraespecífica exerce mais influência.
MacArthur (1958) mostra justamente que a coexistência de cinco espécies de mariquitas (Dendroica sp.) com preferências ecológicas similares só é possível porque α < 1 para todas as espécies, ou seja, a competição entre indivíduos da mesma espécie regula muito mais a população do que a competição com outras espécies. Assim, há uma clara partilha de recursos através da distinção entre o habitat de forrageio dessas aves e, assim, há uma diferenciação de nicho entre as cinco espécies analisadas. Em anos posteriores, David Tilman desenvolve um modelo mecanístico para descrever a competição interespecífica, baseando-se na relação entre os consumidores e seus recursos (Tilman, 1982). Com esse modelo, Tilman demonstra que a coexistência entre competidores é possível quando uma espécie X é melhor competidora do que a espécie Y para um recurso compartilhado “A”, porém Y tem vantagem competitiva sobre X quando se refere a um recurso “B”. Essa última dinâmica descrita representa o que são os tradeoffs associados à habilidade competitiva, que nada mais são do que diferentes performances das espécies frente ao uso de recursos, permitindo a coexistência das mesmas em diferentes escalas (Kneitel & Chase, 2004). Assim, através da dedução de ambos os modelos (fenomenológico e mecanístico) é possível concluir que a coexistência estável entre espécies competidoras é possível quando a dinâmica da competição intraespecífica por determinado recurso é mais forte do que a competição interespecífica (Mittelbach & McGill, 2019).
Desde a conceituação da importância da competição para a dinâmica de uma comunidade biológica houve uma explosão de estudos que investigaram, sendo na maioria das vezes encontrada, a influência da competição interespecífica na estruturação da comunidade (Connel, 1983; Schoener, 1983). A importância da competição foi tão evidenciada que passou a ser incorporada e representar uma das principais formas de investigação dentro de outro conceito bastante relevante na Ecologia de Comunidades, o nicho ecológico das espécies. O nicho de uma espécie, segundo Hutchinson (1957) descreve a amplitude das condições físicas e biológicas (bióticas – incluindo as interações entre espécies, como competição; e abióticas) necessárias para a manutenção da mesma, podendo ser definido como um hipervolume n-dimensional. Mais recentemente, com o avanço das ferramentas analíticas em nível molecular e elementar, novas abordagens vêm sendo empregadas com o intuito de mensurar o nicho de uma espécie, como é o caso do nicho isotópico. Através da análise de isótopos estáveis de carbono (δ13C) e nitrogênio (δ15N) de um indivíduo é possível fazer inferências a respeito do tipo de habitat utilizado pelo mesmo e sobre a sua dieta, respectivamente. De modo que, através da associação bivariada dos valores isotópicos de carbono e nitrogênio, é possível compreender dois importantes eixos que compõem o nicho de uma espécie (Newsome et al., 2007).
A amplitude do nicho isotópico de cinco guildas tróficas (frugívoros, granívoros, insetívoros, nectarívoros e onívoros) de aves da Mata Atlântica sofreu uma redução bastante acentuada (28–70%) no último centenário, muito provavelmente em decorrência dos intensos impactos antropogênicos neste bioma (Navarro et al., 2021a). A expansão humana sobre áreas naturais influenciou diretamente para uma depleção não somente na diversidade de espécies de vertebrados, mas também na riqueza e abundância de invertebrados e plantas, os quais são recursos fundamentais para as aves (Dirzo & Raven, 2003; Haddad et al., 2015; Püttker et al., 2020). Apesar do encolhimento dos nichos das guildas de aves poder ser reflexo direto da redução temporal na disponibilidade e diversidade de recursos, em Navarro et al. (2021a) não foi analisado se a sobreposição de nichos entre as espécies de uma mesma guilda também reduziu proporcionalmente ou se atualmente há maior sobreposição do que era observado no passado. Medir a sobreposição de nichos entre espécies não necessariamente garante que há competição interespecífica por recursos, uma vez que os recursos podem ser abundantes o suficiente para manter as espécies (Morin, 2011). No entanto, conhecendo-se um pouco sobre a história natural das espécies e seus hábitos de forrageio, pode-se especular informações interessantes a respeito da sobreposição de nichos entre espécies de uma guilda trófica e sua possível relação com a dinâmica de competição entre as mesmas. Seguindo na linha da análise de isótopos estáveis, é perfeitamente possível calcular a sobreposição do nicho isotópico através de modelos matemáticos e pacotes estatísticos já existentes (e.g., SIBER no R por Jackson et al., 2011; rKIN no R por Eckrich et al., 2020), uma vez que haja suficiência amostral de indivíduos.
As espécies das guildas analisadas são em sua grande maioria consideradas generalistas e comuns em florestas fragmentadas da Mata Atlântica. Ainda assim, essas aves demonstram que paisagens sob intensos impactos antropogênicos com mudanças no uso do solo, e consequente redução na disponibilidade de recursos e habitat florestal, afetam diretamente os seus nichos (Navarro et al., 2021b). Se realizada a análise de sobreposição de nicho isotópico entre espécies de uma guilda e for encontrada alta sobreposição, pode-se pensar na possibilidade de ocorrência de competição interespecífica por recursos limitados, caso não haja partilha de recursos ou existência de tradeoffs. De modo que, havendo a persistência ou intensificação dos impactos antropogênicos, e consequente redução na disponibilidade e diversidade de recursos, essas aves podem passar a competir mais fortemente, podendo futuramente levar a exclusão competitiva de algumas espécies da guilda que tenham menor habilidade competitiva ou que dependam da existência de maior abundância de recursos. Contrariamente, caso se observe baixa sobreposição de nicho entre espécies de uma mesma guilda, pode-se pensar na hipótese de coexistência estável, já que aqui a competição intraespecífica teria maior influência para a dinâmica da comunidade, o que permitiria essas espécies coexistirem mesmo sob as fortes pressões antropogênicas atuais sobre seus recursos.
Referências bibliográficas
Connell, J. H. (1983). On the prevalence and relative importance of interspecific competition: evidence from field experiments. The American Naturalist, 122(5), 661-696.
Dirzo, R., & Raven, P. H. (2003). Global state of biodiversity and loss. Annual review of Environment and Resources, 28(1), 137-167.
Eckrich, C. A., Albeke, S. E., Flaherty, E. A., Bowyer, R. T., & Ben‐David, M. (2020). rKIN: Kernel‐based method for estimating isotopic niche size and overlap. Journal of Animal Ecology, 89(3), 757-771.
Gause, G. F. (1934). The Struggle for Existence: A Classic of Mathematical Biology and Ecology. Hafner Publishing Company, New York.
Haddad, N. M., Brudvig, L. A., Clobert, J., Davies, K. F., Gonzalez, A., Holt, R. D., … & Townshend, J. R. (2015). Habitat fragmentation and its lasting impact on Earth’s ecosystems. Science advances, 1(2), e1500052.
Hutchinson, G.E. (1957) Concluding remarks. Population Studies: Animal Ecology and Demography. Cold Spring Harbor Symposium on Quantitative Biology, 22, 415–457.
Jackson, A. L., Inger, R., Parnell, A. C., & Bearhop, S. (2011). Comparing isotopic niche widths among and within communities: SIBER–Stable Isotope Bayesian Ellipses in R. Journal of Animal Ecology, 80(3), 595-602.
Kneitel, J. M., & Chase, J. M. (2004). Trade‐offs in community ecology: linking spatial scales and species coexistence. Ecology Letters, 7(1), 69-80.
MacArthur, R. H. (1958). Population ecology of some warblers of northeastern coniferous forests. Ecology, 39(4), 599-619.
Mittelbach, G. G., & McGill, B. J. (2019). Community ecology. Oxford University Press, Oxford.
Morin, P. J. (2011). Community ecology. 2nd ed. Wiley-Blackwell, Hoboken.
Navarro, A. B., Magioli, M., Bogoni, J. A., Silveira, L. F., Moreira, M. Z., Alexandrino, E. R., … & Ferraz, K. M. P. M. D. B. (2021a). Isotopic niches of tropical birds reduced by anthropogenic impacts: a 100‐year perspective. Oikos, 130(11), 1892-1904.
Navarro, A. B., Magioli, M., Bogoni, J. A., Moreira, M. Z., Silveira, L. F., Alexandrino, E. R., … & Ferraz, K. M. P. M. B. (2021b). Human-modified landscapes narrow the isotopic niche of neotropical birds. Oecologia, 196(1), 171-184.
Newsome, S. D., Martinez del Rio, C., Bearhop, S., & Phillips, D. L. (2007). A niche for isotopic ecology. Frontiers in Ecology and the Environment, 5(8), 429-436.
Püttker, T., Crouzeilles, R., Almeida-Gomes, M., Schmoeller, M., Maurenza, D., Alves-Pinto, H., … & Prevedello, J. A. (2020). Indirect effects of habitat loss via habitat fragmentation: A cross-taxa analysis of forest-dependent species. Biological Conservation, 241, 108368.
Schoener, T. W. (1983). Field experiments on interspecific competition. The American Naturalist, 122(2), 240-285.
Tilman, D. (1982). Resource Competition and Community Structure. Princeton University Press, New Jersey.
A importância da integração de processos históricos e regionais na compreensão da montagem de comunidades locais
Rosana Campos Paschoalino
Existem diversos processos ecológicos, históricos e biogeográficos que norteiam a montagem das comunidades ecológicas atuais (Ricklefs 1976). Os processos históricos envolvem eventos ao longo do tempo que determinam a situação atual de uma comunidade em escala local (Cavender-Bares et al. 2009). Esses eventos podem estar relacionados à dinâmica geológica do lugar e às relações filogenéticas das espécies que formaram ao longo do tempo, seja por especiação, seleção, deriva e/ou dispersão (MacArthur 1969; Ricklefs 1987; Vellend 2010). Além da história evolutiva das espécies e das mudanças sofridas no ambiente, os processos ditos inicialmente regionais, estão relacionados ao pool regional de espécies, responsável por gerar e manter a diversidade das comunidades locais (Ricklefs 1987). Isso significa que se uma espécie se extingue dentro da comunidade, ela pode ser ‘renovada’ com a chegada de novas populações da mesma, ou substituída com a chegada de novas espécies do pool regional, já que comunidades ecológicas são abertas e sempre suscetíveis a eventos de migração (Harrison e Cornell 2008).
É de grande conhecimento que processos ecológicos são resultados de processos históricos e regionais, mas pouco se discute e investiga que processos históricos e regionais também podem ser resultantes de processos ecológicos locais (Wiens e Donoghue 2004). Por isso a integração e investigação do passado e presente de comunidades locais devem ser feitas, inclusive em estudos empíricos de ecologia de comunidades, mesmo quando a base de dados é restrita (Harrison e Cornell 2008). Assim, ecólogos devem investigar processos locais (bióticos e abióticos) e padrões que formam a composição e riqueza das comunidades a fim de uma melhor compreensão sobre processos regionais e históricos, como uma via de mão dupla (o que localmente influencia os limites do alcance dos clados envolvidos) (Wiens e Donoghue 2004). Para isso, podemos contar com o avanço no desenvolvimento de ferramentas filogenéticas, surgindo um novo campo da filogenia para ecologia de comunidades (Webb et al. 2002; Cavender-Bares et al. 2009), além de modelos matemáticos desenvolvidos que incorporam cada vez mais características ambientais, relações bióticas, história evolutiva, tudo num mesmo ‘balaio’ a fim de inferir processos e padrões ecológicos de uma maneira mais integrativa (Johnson e Omland 2004).
Focando agora nos processos regionais, sabemos também da forte influência dos mesmos na estruturação de comunidades locais. Cornell em 1985, estudou espécies de carvalhos e vespas-da-galha e encontrou que o tamanho do alcance regional dos carvalhos prediz a riqueza regional total e a riqueza local dos carvalhos e das vespas que ocupam cada espécie de árvore. Uma conclusão particular do trabalho é a importância do estudo ser conduzido em escalas espaciais e temporais bem definidas, de acordo com o(s) grupo(s) taxonômico(s) envolvido(s), para que assim o viés de inferência sobre processos e padrões seja menor. Além disso, devemos considerar que entre regiões e pequenas localidades também existe um sistema de ‘mão dupla’, onde sabemos, como já dito anteriormente, que a composição regional influencia as localidades que a contém. Porém o contrário também é válido, o que acontece em escala local influencia a composição regional, como num sistema de retroalimentação. Mas ainda assim, devemos considerar que a região pode apresentar características independentes e particulares, como a heterogeneidade ambiental e histórico biogeográfico diferente das localidades (Harrison e Cornell 2008). Por exemplo, uma região de savana contém um pool regional que influencia a composição das comunidades em diferentes fitofisionomias dentro da savana e o que acontece dentro das fitofisionomias também influencia a região da savana como um todo. Mas ainda assim, cada fitofisionomia possui um histórico biogeográfico diferente e que a heterogeneidade ambiental (e.g. pluviosidade, cobertura de serapilheira) influencia muito mais a região como um todo do que cada fitofisionomia em particular. Para tentar ilustrar esse cenário, utilizei a figura adaptada de Harrison e colaboradores (2006), onde os autores desenvolvem um modelo para tentar entender as relações e a força/direção da interação de diferentes fatores (ambientais, regionais e locais) na riqueza local de espécies vegetais (Fig. 1).
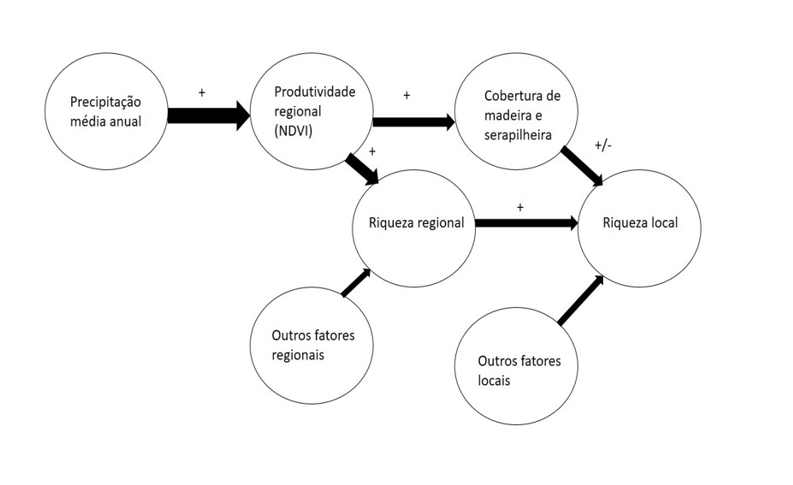 Figura 1- Resultado do modelo de equação estrutural multiescala dos correlatos da riqueza de espécies. A largura das setas indica as forças relativas das vias. Um ‘+’ indica um efeito positivo e um ‘+⁄-‘) indica um efeito unimodal. A figura é adaptada de Harrison et al. 2006 e a legenda é reproduzida de Harrison e Cornell 2008.
Figura 1- Resultado do modelo de equação estrutural multiescala dos correlatos da riqueza de espécies. A largura das setas indica as forças relativas das vias. Um ‘+’ indica um efeito positivo e um ‘+⁄-‘) indica um efeito unimodal. A figura é adaptada de Harrison et al. 2006 e a legenda é reproduzida de Harrison e Cornell 2008.
Segundo a Teoria dos Refúgios, a Amazônia sofreu diversas flutuações em seu clima e vegetação ao longo do tempo, explicando parte da alta biodiversidade endemismo, onde espécies se dispersaram por diversos pontos do bioma e depois ficaram restritas a parte deles e diferentes formações vegetacionais surgiram e desapareceram nesse extenso processo (Haffer 1969; Ab’ Saber 1992). As regiões de savanas estão espalhadas em diversas manchas imersas na matriz de floresta ombrófila do bioma amazônico (Almeida-Júnior 1986). Em Carajás, no sudeste do estado do Pará, elas são encontradas nos topos das montanhas das serras que compreendem a Floresta Nacional de Carajás (Flonaca). O soerguimento das rochas que formam as serras é datado do período Pré-Cambriano e seu assentamento se deu no final do Mesozoico, início do período Terciário, concluindo que são formações rochosas bem antigas (Ab’Saber 1986). Dentro das áreas dessas savanas, encontramos basicamente três tipos de fitofisionomias bem distintos: capões de mata (pequenas ilhas de vegetação estruturalmente mais desenvolvidas), campos brejosos e campos rochosos, que estão uniformemente distribuídas pelas serras (Silva et al. 1996). No meu trabalho de mestrado amostramos comunidades locais de anfíbios anuros, com espécies endêmicas das savanas e outras compartilhadas com a floresta. Inicialmente, eu e meus colaboradores, investigamos a possível divergência de espécies de anuros e seus atributos funcionais entre as diferentes fitofisionomias e posteriormente, incluímos a comparação das comunidades locais entre as diferentes serras da Flonaca.
Pensando na integração de diferentes processos, seria muito interessante investigar possíveis processos históricos e regionais envolvidos (tanto da área de estudo quanto das espécies) e analisar a influência do pool regional sobre a estruturação das comunidades locais. Anfíbios possuem um longo histórico evolutivo na Terra (350 Ma) (Instituto Rã-Bugio 2022), com tempo suficiente para grandes eventos de dispersão e estabilização em vários ambientes. Não seria diferente com a dispersão e estabilização das espécies nessas áreas de savana imersas na floresta amazônica, mas ao mesmo tempo, isso pode ser um impedimento para muitas delas se dispersarem. Outra questão interessante a se levantar é: e como as espécies típicas de outras áreas abertas (e.g. Cerrado) conseguiram chegar nessas ilhas de savanas amazônicas?
A partir do compilado dos atributos funcionais das espécies, a análise da formação de diferentes grupos funcionais pode indicar se essas comunidades se estruturam a partir de relações bióticas (competição) ou por influência do ambiente, do histórico da área ou pelo pool regional, pensando em um modelo semelhante ao de Harrison e colaboradores (2006). Além disso, incluir a filogenia das espécies e dados das características ambientais de cada fitofisionomia, pode gerar o modelo proposto para compreender o grau de influência de cada processo diferente que possa definir os padrões de diversidade de anfíbios nas savanas. Mas pensando no que já temos e para finalizar, apesar de ter sido um estudo experimental, com coleta de dados limitada e pouco enviesada, as informações levantadas fornecerão subsídios para a compreensão da outra via que poucos ecólogos se direcionam: como processos locais podem influenciar processos regionais e até mesmo históricos. E história é o que não falta nas savanas amazônicas de Carajás.
Referências bibliográficas
Ab’Saber, A. (1986) Geomorfologia da região. Em: Carajás: Desafio político, ecologia e desenvolvimento, 1st ed (ed J. Almeida Jr.) pp. 88-124. Brasiliense, São Paulo.
Ab’Sáber, A. (1992) A teoria dos refúgios: Origem e significado. Revista do Instituto Florestal. Edição Especial.
Almeida-Júnior, J. M. G. (1986) Carajás: Desafio político, Ecologia e Desenvolvimento. CNPq, 1st ed. Brasiliense, São Paulo.
Cavender-Bares, J; Kozak, K.H; Fine, P.V; Kembel, S.W. (2009) The merging of community ecology and phylogenetic biology. Ecology letters, doi: 10.1111/j.1461-0248.2009.01314.x.
Haffer, J. (1969) Speciation in Amazonian Forest Birds. Science, New Series, v. 165, n. 3889.
Harrison, S; Safford, H.D; Grace, J.B; Viers, J.H; Davies, K.F. (2006). Regional and local species richness in an insular environment: serpentine plants in California. Ecological Monographs, 76,41–56.
Harrison, S; Cornell, H. (2008) Toward a better understanding of the regional causes of local community richness. Ecology Letters, doi: 10.1111/j.1461-0248.2008.01210.x.
Instituto Rã-bugio. Anfíbios. <http://ra-bugio.org.br/anfibios_sobre_01.php>. Acesso em 09 de Março de 2022. Johnson, J.B; Omland, K.S. (2004) Model selection in ecology and evolution. Trends in Ecology and Evolution, doi.org/10.1016/j.tree.2003.10.013.
MacArthur, R.H. (1969) Patterns of communities in the tropics. Biological Journal of the Linnean Society, Volume 1, Issue 1-2, Pages 19–30, https://doi.org/10.1111/j.1095-8312.1969.tb01809.x.
Ricklefs, R.E. (1976) História, Biogeografia e Diversidade em: A economia da Natureza. 1 ed.
Ricklefs, R.E. (1987) Community Diversity: Relative Roles of Local and Regional Processes. Science 235: 167-171.
Silva, M.F; Secco, R.S; Lobo, M. (1996) Aspectos ecológicos da vegetação Rupestre da Serra dos Carajás (PA). Acta Amazônica 26 (1/2): 17-46.
Vellend, M. (2010) Conceptual synthesis in community ecology. The Quaterly Review of Biology 85: 183-206.
Webb, C.O; Ackerly, D; McPeek, M.A; Donoghue, M.J. (2002) Phylogenies and Community Ecology. Annual Review of Ecology and Systematics, Vol. 33, pp. 475-505.
Wiens, J; Donoghue, M.J. (2004) Historical biogeography, ecology and species richness. Trends in Ecology and Evolution Vol.19 No.12.
Os três pilares do conhecimento e a importância de integra-los
César Santificetur
Com os problemas ambientais da atualidade, o estudo de ecologia acaba tendo um importante papel e também um grande desafio para buscar respostas e estratégias que possam diminuir a degradação do meio ambiente. Para muitos autores este conhecimento ainda é insuficiente, para tornar essas descobertas em ecologia, uma ferramenta concreta que dê resoluções para a crise ambiental atual (Simberloff, 2004; Krebs, 2006). Um dos pontos criticados por esses autores, é a falha na construção de conhecimentos teóricos e empíricos de um modo conjunto. A ecologia de comunidade se interessa em compreender alguns aspectos dos sistemas naturais, sendo eles as características das assembleias, composição das espécies, abundâncias relativas, diversidade e estrutura da cadeia alimentar (Werner 1998). Assim, como muitas ciências, a ecologia de comunidade utiliza abordagens referentes aos três pilares de geração de conhecimento (observacional, experimental e teórico) para tentar solucionar suas questões.
Segundo Tilman (1989) o foco em padrões amplos e repetíveis faz com que possamos inferir algo sobre o sistema ecológico que estamos estudando. A abordagem observacional foca em padrões que se repetem permitindo assim, captar fenômenos que interferem em um sistema ecológico. A presença de um padrão claro e marcante geralmente indica a presença de fortes fatores causadores. Werner (1998) enumera quatro vantagens de uma abordagem observacional em um programa de pesquisa. O primeiro é relacionado ao padrão observado e o quão claro ele é, facilitando assim estudos posteriores que possam surgir a partir da singularidade de um sistema. Ligado a isso a segunda vantagem a capacidade que essa abordagem tem em orientar o desenvolvimento de teorias e direciona-la para trabalhos experimentais a partir de observações e comparações em campo. Além disso, a abordagem observacional tem a capacidade de fazer previsões explícitas em trabalho com a experimental e teórica, fornecendo elementos que ajudam a auto-correção em um programa. A quarta vantagem citada por Werner é a presença de um padrão claro, isso indica fortes causadores ajudando assim a manter a motivação quando há em becos em trabalhos teóricos e experimentais. Schiesari (2016) ainda ressalta que abordagens observacionais são fortalecidas se usadas comparativamente em gradientes ambientais, trazendo ótimos insights sobre fatores que organizam algumas comunidades ecologias. Na década de 1960 e 170 a abordagem observacional sofreu diversas críticas devido ao seu uso excessivo como ferramenta para testar teorias ecologias, porem a sua importância se dá principalmente como uma primeira compreensão de um sistema, gerando uma explicação ou uma resposta alternativa. Com isso, a abordagem observacional ainda pode ser considerada uma ferramenta importante para um projeto de pesquisa.
A abordagem experimental pode ser definida como um conjunto de procedimentos estabelecidos para verificar se uma variável interfere ou não em um sistema ecológico. Um experimento pode em suma controlar todas as variáveis de um sistema, menos a que se deseja medir. Em uma abordagem experimental existe uma grande preocupação para que haja um certo equilíbrio entre o controle total das variáveis para assim poder detectar causa-e-efeito e o quando essa manipulação afasta o sistema do que realmente acontece na natureza. Não há dúvidas que a abordagem experimental fornece uma das melhores ferramentas para atingir esses objetivos em pesquisas ecologicas (Schoener, 1983), porém, muitas vezes esse tipo de estudo é criticado por sua falta de generalidade, escala especial e temporal (Carpenter, 1996; Schindler, 1998). A modelagem fornece outra maneira de decifrar informações ecológicas, porem essa ferramenta depende muito de modelos matemáticos. Atualmente a modelagem leva a uma melhor compreensão dos processos através de abordagens interdisciplinares (Bashkin, 2002), com isso, um grande foco nas ciências ecológicas é a produção de abordagens quantitativas testáveis experimentalmente (Loreau, 2010).
Enquanto cientistas observacionais e experimentais estudam sistemas reais tanto em laboratório como em campo, cientistas teóricos estudam sistemas idealizados (Wilson, 2000), tal representação teórica é denominada modelo. Existem diversos exemplos de modelos aplicados em diversas ciências, em ecologia pode-se ressaltar os de dinâmica de populações (Gotelli, 2007) e comunidades (May, 1975). A abordagem teoria utiliza a observação e a experimentação para incorpora generalização em um programa de pesquisa. Essa abordagem faz com que os pesquisadores façam suposições claras e inequívocas, para que haja coerência nos fatos, e assim, esclareça hipóteses, identifique elementos-chave em um sistema, e examine as consequências lógicas de novas ideias. A abordagem teórica, portanto, tem a vantagem de esclarecer pensamentos sobre um fenômeno, além disso, a teoria é útil como guia para padronizar dados permitindo assim sua comparação mesmo em diferentes sistemas. Em suma, a abordagem teórica tem como objetivo expor, em forma de leis, como fenômenos naturais podem funcionar e diferente das outras das outras, o produto a abordagem teórica não é a observação de padrões ou resultado de experimentos, e sim ideias (Mandai, 2014). A abordagem teórica em um programa de pesquisa acaba necessariamente aumentando a sua generalidade, porem a falta de uma conversa construtiva entre a parte teórica e a empírica nas pesquisas limitam os avanços de ambos. Kareiva (1989) defende que mais esforços sejam dedicados à elaboração de modelos que incluam parâmetros concretos e diretamente mensuráveis, mesmo que isso leve a modelos menos gerais, mas que assim possam receber mais atenção de outros pesquisadores. Com tudo isso, Cruz et al. (2007) sugere que ainda há um hiato entre as produções empíricas e teóricas, isso faz com que o conhecimento ecológico se dê sem um aprofundamento teórico.
Werner (1998), na conclusão do seu trabalho, mostra a importância da integração destas abordagens em um programa de pesquisa. Para que isso ocorra, devemos estar cientes dos pontos fracos e fortes de cada abordagem, e assim, possamos extrair o melhor de cada uma delas, além de ter uma perspectiva mais equilibrada sobre as questões ecológicas. Apesar de ser um trabalho de 1998 essa problemática parece ser ainda bastante atual. Cruz et al. (2007) ainda sugere esforços na pesquisa histórica e epistemológica. Com isso, o entendimento das abordagens utilizadas em ecologia de comunidade me faz entender melhor a pesquisa de ecologia como um todo. Minha pesquisa, tem uma abordagem observacional clara, entender todas as abordagens fizeram com que eu entendesse suas limitações e também o papel que ela tem em um dentro da ecologia de comunidade como um todo. Assim posso também focar em dar subsídios para as outras abordagens. Outro ponto importante do entendimento de outras abordagens foram as procuras de artigos experimentais e teóricas que se relacionam com o tema da minha pesquisa, me dando mais suporte teórico sobre o tema. Por fim, a frase escrita por Lawrence Slobodkin resume bem a importância de se integrar às três abordagens, e com certeza me norteará em pesquisas futuras. “… de certa forma a distinção entre um ecólogo teórico, um de laboratório e um de campo, é que o teórico lida com todos os mundos concebíveis enquanto o ecólogo de laboratório lida com todos os mundos possíveis e o ecólogo de campo está confinado ao mundo real. O ecólogo de laboratório precisa perguntar ao teórico se o mundo possível é um mundo interessante, e perguntar ao ecólogo de campo se este mundo é relacionado de alguma forma com o mundo real”.
Referências bibliográficas
Bashkin, V. N. 2002. Modern Biogeochemistry. Boston, MA: Kluwer Academic Publishers.
Carpenter, S. R. 1996. Microcosm experiments have limited relevance for community and ecosystem ecology. Ecology 77, 677–680.
Cruz, L.; Rocha, P.; El-Hani, C. 2007. A prática científica na ecologia de comunidades: diálogos entre empirismo e teoría na literatura científica. Filosofia e História da Biologia, v. 2, n. 1, p. 257-278.
Gotelli NJ. 2007. Ecologia. Londrina: Editora Planta.
Kareiva, P. 1989. Renewing the dialog between theory and experiments in population ecology. pp. 68-88. in J. Roughgarden, R. M. May, and S. A. Levin, eds. Perspectives in ecological theory. Princeton University Press. 394pp.
Krebs, C. J. 2006. Ecology after 100 years: progress and pseudoprogress. New Zealand Journal of Ecology 30 (1): 3-11.
Loreau, M. 2010. From Populations to Ecosystems: Theoretical Foundations for a New Ecological Synthesis. Princeton, NJ: Princeton University Press.
Mandai, C. Y. 2014. Abordagem teórica na ecologia: uma visão do mundo através de modelos. Revista da Biologia.
May RM. 1975. Patterns of species abundance and diversity. In: Cody ML, Diamond JM, editors. Cambridge: Harvard University Press p81-120.
Schiesari, Luis. Perspectivas Metodológicas. Ecologia Teórica e Aplicada em Ambientes Modificados. 2016. Disponível em: https://lschiesari.wixsite.com/ecologia-aplicada. Acesso em: 03/03/2022.
Schindler, D. W. 1998. Replication versus realism: the need for ecosystem-scale experiments. Ecosystems 1, 323–334.
Schoener, T. W. 1983. Field experiments on interspecific competition. Am. Nat. 122, 240–285.
Simberloff, D. 2004. Community ecology: is it time to move on? The American Naturalist 163 (6): 787-799.
Tilman, D. 1989. Discussion: Population dynamics and species interactions. pp 89-101. in J. Roughgarden, R. M. May, and S. A. Levin, eds. Perspectives in ecological theory. Princeton University Press. 394pp.
Werner, E. E. 1998. Ecological experiments and a research program in community ecology. Experimental ecology: issues and perspectives, 3-26.
Wilson W. 2000. Simulating ecological and evolutionary systems in C. Cambridge: Cambridge University Press
Planta invasora como potencial força geradora de estados alternativos estáveis de equilíbrio
Marianela Pini
Na natureza, existem sistemas que são dinâmicos, ou seja, que seu estado pode ser compreendido como um balanço de processos. Esse balanço leva o sistema ao equilíbrio, e se o sistema tende a voltar a esse ponto de equilíbrio após uma perturbação, ele é considerado como estável. Um clássico exemplo é uma população hipotética com taxas de nascimento e mortes que chegam ao mesmo valor em um ponto de equilíbrio, que é a capacidade de carga.
Os ecossistemas nem sempre estão em equilíbrio porque as condições ambientais mudam, e podem responder de diferentes maneiras a essas mudanças. Em sistemas dinâmicos, essa resposta pode ser gradativa ou de forma rápida se atinge um valor crítico da condição dada (figura 1a e b). Um terceiro caso é apresentar mais de um estado de equilíbrio possível em um mesmo conjunto de condições, chamados de estados alternativos estáveis (Scheffer & Carpenter, 2003). Nesse sentido, o sistema pode apresentar dois estados de equilíbrio estáveis separados por um estado instável, como aparece na figura 1c. Observando na figura, se o sistema se encontra no ramo superior e a condição vai aumentando, o estado não muda, mas vai chegando no ponto crítico (f2), onde uma leve mudança ocasiona a transição abrupta para o estado inferior, chamado de “catástrofe”. Quanto mais perto do ponto crítico menor é a resiliência, ou seja, a capacidade do sistema de permanecer no mesmo estado, e assim ocorre essa transição brusca (May, 1977; Scheffer et al., 2001).
 Figura 1. Representação esquemática de como o estado de equilíbrio de um ecossistema pode variar dependendo das condições (Scheffer, 2009). A figura a e b são sistemas com um equilíbrio estável e a figura c representa um sistema com dois equilíbrios estáveis (linhas cheias) e um instável (linha tracejada). As setas representam a direção do sistema quando não está em equilíbrio.
Figura 1. Representação esquemática de como o estado de equilíbrio de um ecossistema pode variar dependendo das condições (Scheffer, 2009). A figura a e b são sistemas com um equilíbrio estável e a figura c representa um sistema com dois equilíbrios estáveis (linhas cheias) e um instável (linha tracejada). As setas representam a direção do sistema quando não está em equilíbrio.
Em resumo, quando as condições mudam, o estado instável fica cada vez mais perto do estado estável até colidir e mudar de estado. Para voltar ao estado original, não basta voltar apenas para as condições originais, é necessário recuar ainda mais nas condições para chegar ao equilíbrio original. Esse processo é chamado de histerese.
O conceito de estados alternativos estáveis se insere no contexto da força de seleção como uma das quatro forças estruturadoras de comunidades segundo Vellend (2010). A seleção é a diferença de fitness entre indivíduos de diferentes espécies e existem processos que beneficiam uma espécie sobre outra. No caso de estados alternativos de equilíbrio, uma mudança no ambiente pode atuar como força de seleção mudando a composição das espécies. Como consequência, a seleção de determinadas espécies permite a coexistência em vários estados de equilíbrio estáveis. Podemos concluir que a teoria de estados múltiplos faz parte da teoria de montagem de comunidades através do nicho, teoria difundida por várias décadas na Ecologia (Chase & Leibold, 2003; Hutchinson, 1957). Nessa teoria, a seleção atua e existe um equilíbrio nas comunidades, onde o sistema é determinístico, ou seja, é possível predizer o seu comportamento.
Os mecanismos que geram estados estáveis alternativos ocasionam sistemas de feedback positivo, que levam a uma escolha de um estado estável ou outro. Esses feedbacks positivos são comuns na natureza e devem ser o suficientemente fortes para que mantenham os estados alternativos estáveis (DeAngelis et al., 1986). Um dos mecanismos é a facilitação. Nesse sentido, uma espécie pode modificar as condições do seu entorno e se beneficiar, ou ajudar outras espécies. Em comunidades de plantas terrestres de ambientes áridos que apresentam dois estados estáveis, um com áreas abertas onde só sobrevivem ervas tolerantes a muita luz e aridez e outro estado estável com vegetação arbórea e condições mais amenas, existe o mecanismo de facilitação (Rietkerk & van de Koppel, 1997). As árvores podem modificar a disponibilidade de água no solo, acumulando matéria orgânica morta, aumentando a infiltração no momento da chuva e disponibilizando maior quantidade de água para as espécies. Além disso, podem gerar sombreamento, diminuindo a temperatura e aumentando a umidade do solo, beneficiando o estabelecimento de plântulas em ambientes onde sem o sombreamento não seria possível (Bertness & Yeh Su Ming, 1994; Stachowicz, 2001). Esses mecanismos ajudam a manter o estado estável de vegetação arbórea. Outro exemplo de facilitação é em escala regional, onde plantas aumentam a evapotranspiração, levando a um aumento de chuvas beneficiando o crescimento da vegetação (Oyama & Nobre, 2003).
No meu projeto de mestrado o objeto de estudo é a planta leguminosa invasora Leucaena leucocephala, que é de grande importância nos ecossistemas, dado que modifica os ambientes e tem efeito sobre as espécies nativas (Aldava Navarro et al., 2017). Diferentes estratégias funcionais da planta invasora poderiam explicar potenciais mecanismos de facilitação para a geração de estados alternativos estáveis. Uma delas é a capacidade de fixar nitrogênio. Com isso, a espécie pode facilitar a disponibilidade de nitrogênio das plantas vizinhas, mas dado que ela cresce rapidamente e forma aglomerações, é possível que o nitrogênio se acumule em grandes quantidades beneficiando só aquelas espécies que toleram esse nível do nutriente e são melhores competidoras. Consequentemente, essas espécies dominam sobre as que não toleram esses níveis da condição, reduzindo a biodiversidade. Esse efeito já foi estudado em outras plantas invasoras fixadoras (Benesperi et al., 2012). Para Leucaena leucocephala, a presença de menos espécies gera menor competição com as plantas nativas e, portanto, uma resposta de retroalimentação positiva, porque ao reduzir a competição aumenta o crescimento da planta invasora. Seria possível então a presença de um estado estável com dominância da planta invasora e grandes quantidades de nitrogênio e outro estado estável sem a espécie invasora com maior biodiversidade. Para voltar ao estado estável original sem Leucaena leucocephala, não bastaria só retirar alguns indivíduos da espécie porque ela também modifica as condições do solo.
Atributos funcionais da espécie invasora como grande quantidade de produção de sementes, curto tempo pré-reprodutivo e crescimento rápido permitem que as populações cresçam de forma rápida (Costa & Durigan, 2010; Hughes, 1998). Dessa maneira, é possível manter o estado alternativo estável onde a espécie domina sobre outras espécies nativas.
Outra estratégia funcional possível que garante sua manutenção é o sombreamento pela copa da espécie. Como foi explicado no exemplo de ambientes áridos, e sabendo que Leucaena leucocephala é encontrada nesses ambientes, o sombreamento pode facilitar o crescimento das plântulas da espécie, gerando melhores condições ambientais. Além disso, se colonizar áreas abertas com plantas com altos requerimentos de luz, como ervas, a espécie invasora provocaria a supressão dessas espécies menos tolerantes à sombra e isso resultaria em uma mudança na fisionomia das espécies da comunidade. Já foi encontrado que essas mudanças reduzem a diversidade, e com isso aumenta a dominância e permanência da espécie invasora (Mello, 2014). Porém, como Leucaena leucocephala pode crescer em áreas degradadas, em vez de exclusão competitiva das espécies nativas pela planta invasora, as plantas nativas podem ser afetadas pelas modificações do habitat e isso leva a uma redução nas suas populações. Portanto, outros fatores além da presença da planta invasora poderiam afetar a exclusão competitiva das espécies nativas. Contudo, a planta leguminosa invasora apresenta potenciais características para ser uma força geradora de estados alternativos estáveis.
Considerando que a força da planta invasora que geraria os estados múltiplos é forte, e dadas as evidências da dominância da espécie e modificação do ambiente, o estado estável do sistema onde está presente Leucaena leucocephala poderia ser considerado como um ecossistema novo (Hobbs et al., 2006). A ideia de ecossistemas novos vem sendo difundida nos últimos anos na Ecologia. Nesse sentido, as abundâncias relativas das espécies são novas com a introdução da invasora, e antes da invasão não existia tal combinação de espécies. Um estudo em Porto Rico mostra como a invasão de espécies exóticas, uma delas, L. leucocephala, muda a composição das florestas, criando novos ecossistemas com dominância de poucas espécies e ambientes mais homogêneos (Colón & Lugo, 2006).
Introduzindo novas espécies o ambiente biótico é modificado e possivelmente reduz o potencial de voltar ao ambiente original. Portanto, são possíveis dois cenários: um sistema com estados alternativos estáveis e a planta invasora atuando como força geradora desses estados, ou um ecossistema novo onde a força de invasão é forte suficiente para mudar o sistema totalmente criando novas combinações de espécies, interações e condições, e o retorno ao estado original é quase impossível (Hobbs et al., 2009). Em qualquer desses cenários, seria plausível explorar melhor sobre as características da espécie invasora para desenvolver futuras ações de manejo eficientes que não desperdicem recursos e aumente a diversidade de espécies nos ecossistemas.
Referências bibliográficas
Aldava Navarro, J., Casanova Lugo, F., Díaz Echeverría, V. F., Escobedo Cabrera, A., Estrada Medina, H., Cetzal IX, W., & Basu, S. K. (2017). Influence of Leucaena leucocephala (Lam.) de Wit (Fabaceae) on the forage yield and forage quality of tropical grasses Brachiaria brizantha (Hochst. ex A. Rich.) Stapf and Panicum maximum Jacq. (Poaceae). International Journal on Agricultural Sciences, 8(2), 133–137.
Bertness, M. D., & Yeh Su Ming. (1994). Cooperative and competitive interactions in the recruitment of marsh elders. Ecology, 75(8), 2416–2429. https://doi.org/10.2307/1940895
Chase, J. M., & Leibold, M. A. (2003). Ecological Niches: Linking Classical and Contemporary Approaches. Chicago University Press.
Colón, S. M., & Lugo, A. E. (2006). Recovery of a subtropical dry forest after abandonment of different land uses. Biotropica, 38(3), 354–364. https://doi.org/10.1111/j.1744-7429.2006.00159.x
Costa, J. N. M. N. da, & Durigan, G. (2010). Leucaena leucocephala (Lam.) de Wit (Fabaceae): Invasive or ruderal? Revista Arvore, 34(5), 825–833. https://doi.org/10.1590/s0100-67622010000500008
DeAngelis, D. L., Post, W. M., & C.Travis, C. (1986). Positive feedback in Natural Systems (S.A. Levin (ed.); Springer V, Vol. 15).
Hobbs, R. J., Arico, S., Aronson, J., Baron, J. S., Bridgewater, P., Cramer, V. A., Epstein, P. R., Ewel, J. J., Klink, C. A., Lugo, A. E., Norton, D., Ojima, D., Richardson, D. M., Sanderson, E. W., Valladares, F., Vilà, M., Zamora, R., & Zobel, M. (2006). Novel ecosystems: Theoretical and management aspects of the new ecological world order. Global Ecology and Biogeography, 15(1), 1–7. https://doi.org/10.1111/j.1466-822X.2006.00212.x
Hobbs, R. J., Higgs, E., & Harris, J. A. (2009). Novel ecosystems: implications for conservation and restoration. Trends in Ecology and Evolution, 24(11), 599–605. https://doi.org/10.1016/j.tree.2009.05.012
Hughes, C. (1998). Monograph of Leucaena(Leguminosae-Mimosoideae). Systematic Botany Monographs, 55, 1–244.
Hutchinson, G. E. (1957). Concluding remarks. Cold Spring Harbor Symposia on Quantitative Biology, 22, 415–427. https://doi.org/10.1201/9781315366746
May, R. M. (1977). Thresholds and breakpoints in ecosystem with a multiplicity of stable states. Nature, 269, 471–477.
Mello, T. J. (2014). Biological invasion in oceanic islands: the case of Leucaena leucocephala (Leguminosae) in Fernando de Noronha. Universidade de São Paulo.
Oyama, M. D., & Nobre, C. A. (2003). A new climate-vegetation equilibrium state for Tropical South America. Geophysical Research Letters, 30(23), 10–13. https://doi.org/10.1029/2003GL018600
Rietkerk, M., & van de Koppel, J. (1997). Alternate Stable States and Threshold Effects in Semi-Arid Grazing Systems. Oikos, 79(1), 69. https://doi.org/10.2307/3546091
Scheffer, M. (2009). Critical Transitions in Nature and Society (Simon A. Levin & S. H. Strogatz (eds.); Princeton).
Scheffer, M., Carpenter, S., Foley, J. A., Folke, C., & Walker, B. (2001). Catastrophic shifts in ecosystems. Nature, 413(6856), 591–596. https://doi.org/10.1038/35098000
Scheffer, M., & Carpenter, S. R. (2003). Catastrophic regime shifts in ecosystems: Linking theory to observation. Trends in Ecology and Evolution, 18(12), 648–656. https://doi.org/10.1016/j.tree.2003.09.002
Stachowicz, J. J. (2001). Mutualism, facilitation, and the structure of ecological communities. BioScience, 51(3), 235–246. https://doi.org/10.1641/0006-3568(2001)051[0235:MFATSO]2.0.CO;2
Vellend, M. (2010). Conceptual Synthesis in Community Ecology. The Quarterly Review of Biology, 85(2), 183–206. https://doi.org/10.1017/CBO9781107415324.004
Restauração ecológica como transição entre estados estáveis alternativos
Joyce Fernandes Ferreira
Na segunda metade do século passado emergiu-se a ideia de que sistemas ecológicos pudessem ocorrer como estados estáveis alternativos (May, 1977), sendo que estados estáveis resultam de um balanço de processos que geram equilíbrios estáveis (Scheffer, 2009). Um exemplo de estado estável pode ser dado por uma população hipotética que tem a densidade regulada pelo equilíbrio das taxas de nascimento e mortalidade (figura 1a) (Scheffer, 2009). Nesse caso, o equilíbrio estável é atingido na capacidade de suporte (K) da população, na qual as taxas de natalidade e mortalidade se equivalem (figura 1a). Assim, esse sistema pode ser representado por uma bola em uma bacia de atração (figura 1b), onde mesmo com perturbações ela retornará ao fundo da bacia. Em sistemas com estados estáveis alternativos existe mais de uma bacia de atração (Scheffer, 2009). Um exemplo de tais sistemas pode ser dado por populações com Efeito Allee, ou seja, que em densidades baixas a taxa de mortalidade da população é maior que a de natalidade (figura 2a). Nesse caso, existe um ponto onde as taxas de mortalidade e natalidade se igualam que se trata de um ponto de equilíbrio instável, sendo o limiar a partir do qual a população tenderá aos estados estáveis de densidade 0 ou K (figura 2a). Nesse sistema, para que haja uma mudança de um estado para outro, a bola tem que sofrer uma perturbação grande o suficiente para fazê-la passar de uma bacia de atração para outra (figura 2b) (Beisner et al., 2003; Scheffer, 2009). Assim, o tamanho da bacia de atração pode ser definida como a resiliência de um estado, que também pode ser definida como a perturbação máxima que pode ser exercida sobre um sistema sem que ele mude para um estado estável alternativo (Scheffer, 2009).
 Figura 1: (a) População hipotética que tem sua densidade definida pelo balanço das taxas de natalidade e mortalidade. Essas taxas se encontram no ponto onde a densidade está na capacidade de suporte (k), um ponto de equilíbrio estável. Nesse ponto, caso a população aumente, a taxa de mortalidade passará a ser maior que a de natalidade, retornando a população a K; caso a população diminua, o inverso ocorrerá e a população também retornará a K. (b) Modelo de paisagem de densidade representando um sistema com um único estado estável (vale ou fundo da bacia de atração) para o qual a bolinha sempre retornará, mesmo que sofra perturbações. Figura adaptada de: Scheffer 2009.
Figura 1: (a) População hipotética que tem sua densidade definida pelo balanço das taxas de natalidade e mortalidade. Essas taxas se encontram no ponto onde a densidade está na capacidade de suporte (k), um ponto de equilíbrio estável. Nesse ponto, caso a população aumente, a taxa de mortalidade passará a ser maior que a de natalidade, retornando a população a K; caso a população diminua, o inverso ocorrerá e a população também retornará a K. (b) Modelo de paisagem de densidade representando um sistema com um único estado estável (vale ou fundo da bacia de atração) para o qual a bolinha sempre retornará, mesmo que sofra perturbações. Figura adaptada de: Scheffer 2009.
 Figura 2: (a) População hipotética que tem sua densidade definida pelo balanço das taxas de natalidade e mortalidade com Efeito Allee, que faz com que, em densidades baixas, a taxa de mortalidade seja maior que a de natalidade. Dessa forma, existem dois estados estáveis e um ponto de equilíbrio instável (representado pelo círculo aberto onde as taxas de natalidade e mortalidade se encontram pela primeira vez, à esquerda), no qual qualquer perturbação pode fazer com que a população vá para um dos estados estáveis de densidade populacional: 0 ou K. (b) Modelo de paisagem de densidade representando um sistema com dois estados estáveis (duas bacias de atração) e um instável (crista ou borda da bacia de atração). No ponto instável, onde se encontra a bolinha branca, qualquer perturbação pode fazer com ela seja atraída para o fundo de uma das bacias de atração. Figura adaptada de: Scheffer 2009.
Figura 2: (a) População hipotética que tem sua densidade definida pelo balanço das taxas de natalidade e mortalidade com Efeito Allee, que faz com que, em densidades baixas, a taxa de mortalidade seja maior que a de natalidade. Dessa forma, existem dois estados estáveis e um ponto de equilíbrio instável (representado pelo círculo aberto onde as taxas de natalidade e mortalidade se encontram pela primeira vez, à esquerda), no qual qualquer perturbação pode fazer com que a população vá para um dos estados estáveis de densidade populacional: 0 ou K. (b) Modelo de paisagem de densidade representando um sistema com dois estados estáveis (duas bacias de atração) e um instável (crista ou borda da bacia de atração). No ponto instável, onde se encontra a bolinha branca, qualquer perturbação pode fazer com ela seja atraída para o fundo de uma das bacias de atração. Figura adaptada de: Scheffer 2009.
Feedbacks positivos são peças necessárias - mas não suficientes - para a existência de estados estáveis alternativos (Kéfi et al., 2016; Scheffer, 2009) e um mecanismo ecológico que comumente está por trás desses feedbacks positivos são as interações de facilitação (Scheffer, 2009). Feedbacks positivos ocorrem quando o resultado das relações ecológicas entre componentes bióticos e abióticos de um sistema promove a manutenção do seu estado (Kéfi et al., 2016). Um exemplo clássico de sistema com estados estáveis alternativos ocorre em lagos rasos que podem apresentar um estado com águas límpidas ou com águas túrbidas (Blindow et al., 1993; Scheffer et al., 1993). O estado de águas límpidas permite que a luz atinja maiores profundidades na coluna d’água, permitindo o estabelecimento de plantas submersas que, por sua vez, mantêm o estado da água límpida por uma série de mecanismos. Um aumento de nutrientes no lago, pode fazer com que o lago mude para o estado de águas túrbidas, aumentando a densidade de fitoplâncton e impedindo o estabelecimento da vegetação submersa.
Além de perturbações, variações nas condições externas ao sistema podem levar a mudanças entre estados estáveis alternativos (Beisner et al., 2003). Isso porque essas condições externas podem alterar a estabilidade do sistema, afetando a resiliência e possibilitando que exista apenas um estado estável ou estados estáveis alternativos (figura 3). Assim, a resposta do sistema às condições externas se apresenta na forma de uma curva dobrada (figura 3) em que um estado pode ser insensível a mudanças externas, até que elas ultrapassem um limite, resultando em uma transição crítica de estado (mudança catastrófica) (Scheffer, 2009). Uma característica importante de estados estáveis alternativos é a histerese (Beisner et al., 2003; Scheffer, 2009), fenômeno que faz com que, para reverter uma mudança catastrófica, seja necessário retornar as condições a níveis mais inferiores do que aquele no qual a mudança ocorreu (figura 3). Esse efeito das condições externas sobre o sistema pode fazer com que variações pequenas nas condições ou pequenas perturbações causem mudanças catastróficas de estados, sendo que o sistema pode não apresentar sinais de que está próximo de uma mudança catastrófica até que ela ocorra.
 Figura 3: Representação esquemática da resposta de um sistema a mudanças nas condições externas. As curvas contínuas representam dois estados de equilíbrio estável diferentes. Se o sistema se encontrar fora dessas curvas ele tenderá a retornar para um dos estados estáveis de acordo com a direção das setas. Se o sistema se encontrar no estado de equilíbrio estável superior e as condições ultrapassarem o ponto F2, haverá uma mudança catastrófica para o outro estado de equilíbrio estável. Porém, para retornar ao estado inicial, as condições deverão ser revertidas a níveis bem mais inferiores do que aquele no qual a mudança ocorreu, ou seja, deverão ultrapassar o ponto F1, fenômeno chamado de histerese. Figura adaptada de: Scheffer 2009.
Figura 3: Representação esquemática da resposta de um sistema a mudanças nas condições externas. As curvas contínuas representam dois estados de equilíbrio estável diferentes. Se o sistema se encontrar fora dessas curvas ele tenderá a retornar para um dos estados estáveis de acordo com a direção das setas. Se o sistema se encontrar no estado de equilíbrio estável superior e as condições ultrapassarem o ponto F2, haverá uma mudança catastrófica para o outro estado de equilíbrio estável. Porém, para retornar ao estado inicial, as condições deverão ser revertidas a níveis bem mais inferiores do que aquele no qual a mudança ocorreu, ou seja, deverão ultrapassar o ponto F1, fenômeno chamado de histerese. Figura adaptada de: Scheffer 2009.
A possibilidade de sistemas ecológicos apresentarem estados estáveis alternativos e de sofrerem mudanças catastróficas difíceis de serem previstas e revertidas pode ter um impacto sobre o manejo de ecossistemas e comunidades (Folke et al., 2002; Suding et al., 2004). O conhecimento sobre os possíveis estados alternativos e sobre os mecanismos que os regulam pode impulsionar estratégias de manejo com objetivos de evitar transições críticas de estados (Scheffer, 2009). Pode-se, por exemplo, criar cenários para visualizar possíveis futuros alternativos e o que pode desencadeá-los, assim como fazer estimativas de risco acerca de tomadas de decisões sobre determinado sistema. Dessa forma, estratégias de manejo podem ser feitas visando aumentar a resiliência de um sistema e diminuir a chance de uma transição crítica (Folke et al., 2002; Scheffer, 2009). Além disso, estratégias de manejo podem ter o objetivo de promover transições críticas de estado, por exemplo, promovendo a transição de um estado degradado para um restaurado (Scheffer, 2009; Suding et al., 2004). Uma maneira de fazer isso é realizando perturbações que impulsionam a mudança de estado em situações em que a resiliência do estado degradado é menor (Scheffer, 2009). Pode-se, por exemplo, aproveitar períodos de El Niño para restaurar ecossistemas de climas secos, pois a maior precipitação causada pelo El Niño poderia diminuir a resiliência do sistema degradado, facilitando o estabelecimento da vegetação que se pretende restaurar (Scheffer, 2009).
Muitos projetos de restauração não são bem-sucedidos em atingir todos seus objetivos (Lockwood & Pimm, 1999), e modelos de estados estáveis alternativos podem explicar o porquê de sistemas degradados poderem ser resilientes à restauração (Suding et al., 2004). Algumas estratégias de restauração são focadas em restabelecer as condições abióticas prévias à mudança de regime, para assim promover um retorno natural da vegetação (Suding et al., 2004). Em algumas situações essas estratégias podem ser bem-sucedidas, porém, em outras, é necessário que ações ativas de manejo sejam realizadas, sugerindo que a situação degradada possa se tratar de um estado estável alternativo (Suding et al., 2004). Dessa forma, seria então necessária uma quebra de seus mecanismos de feedbacks para que haja uma mudança de estado. Dentre os mecanismos de feedbacks que podem aumentar a resiliência de sistemas degradados estão os efeitos gerados pelas espécies presentes no sistema degradado, que podem mudar as características do ecossistema beneficiando a si mesmas (Suding et al., 2004). Além disso, em sistemas degradados, a fragmentação e perda de conectividade na paisagem pode limitar a recolonização do sistema por espécies nativas, também aumentando a resiliência do sistema degradado (Suding et al., 2004).
De acordo com a hipótese do gradiente de estresse, interações de facilitação podem se tornar mais importantes em condições mais severas (Bertness & Callaway, 1994), como pode ser o caso de ambientes degradados, por terem condições muito diferentes das condições da vegetação original. Dessa forma, a facilitação entre plantas pode ser um mecanismo importante para o estabelecimento da comunidade em restaurações ecológicas (Padilla & Pugnaire, 2006), nas quais a presença de um indivíduo pode melhorar as condições microclimáticas e edáficas (Callaway, 1995) e favorecer o desenvolvimento de plantas vizinhas (Bueno & Llambí, 2015; Gómez-Aparicio et al., 2004). Interações de facilitação podem atuar como componentes de feedbacks positivos de estados estáveis (Kéfi et al., 2016), podendo então beneficiar a transição do estado degradado para o restaurado. Assim, projetos de restauração podem se beneficiar de mecanismos de facilitação (Padilla & Pugnaire, 2006) e contar com métodos com o potencial de promovê-los, como plantios em que as espécies são plantadas de forma agrupada, com menor espaçamento entre si (Silliman et al., 2015; Corbin & Holl, 2012).
Entretanto, interações de competição também podem ocorrer entre plantas vizinhas, de forma que o efeito de vizinhança entre elas seja dado pelo saldo entre as interações de competição e facilitação (Callaway & Walker, 1997), que pode ser positivo ou negativo. Além disso, os efeitos de vizinhança podem ser influenciados pelas condições ambientais, pelas características das espécies e pelo estágio ontogenético dos indivíduos envolvidos (Armas & Pugnaire, 2009; Callaway & Walker, 1997; Miriti, 2006). Assim, penso que o resultado da dinâmica de feedback, além de depender do sinal e intensidade dos diferentes efeitos de vizinhança entre os indivíduos presentes no sistema (Kéfi et al., 2016), pode variar de acordo com as espécies escolhidas para a restauração ou mesmo com o passar do tempo.
Em meu projeto de mestrado vamos trabalhar com dados do monitoramento de um plantio de restauração de floresta de restinga que contou com um delineamento experimental com dois tratamentos. Em um deles as mudas foram plantadas em agrupamentos multiespecíficos incluindo espécies pioneiras e não pioneiras e, no outro, mudas das mesmas espécies foram plantadas isoladamente. Temos como objetivo avaliar os efeitos de vizinhança entre as espécies que estão nos agrupamentos multiespecíficos, considerando os grupos sucessionais das espécies e o tempo transcorrido desde o início do plantio. Assim, acredito que explorar como os efeitos de vizinhança podem ser influenciados pela identidade das espécies e como podem variar ao longo do tempo, pode fornecer indicativos sobre a possibilidade da ocorrência de mecanismos de feedbacks positivos nesse tipo de plantio, que poderiam então beneficiar a transição de um estado degradado para um restaurado.
Referências bibliográficas
Armas, C., & Pugnaire F. I. (2009). Ontogenetic shifts in interactions of two dominant shrub species in a semi-arid coastal sand dune system. Journal of Vegetation Science, 20(3), 535–546
Beisner, B. E., Haydon, D. T., & Cuddington, K. (2003). Alternative stable states in ecology. Frontiers in Ecology and the Environment, 1(7), 376-382.
Bertness, M. D., & Callaway, R. (1994). Positive interactions in communities. Trends in Ecology and Evolution, 9(5), 191–193.
Blindow I., Andersson, G., Hargeby, A., & Johansson, S. (1993). Long-term pattern of alternative stable states in two shallow eutrophic lakes. Freshwater Biology, 30(1): 159-167.
Bueno, A., & Llambí, L. D. (2015). Facilitation and edge effects influence vegetation regeneration in old-fields at the tropical Andean forest line. Applied Vegetation Science, 18(4), 613–623.
Callaway, R. M. (1995). Positive interactions among plants. The Botanical Review, 61(4), 306–349.
Callaway, R. M., & Walker, L. R. (1997). Competition and facilitation: A synthetic approach to interactions in plant communities. Ecology, 78(7), 1958–1965.
Corbin, J. D., & Holl, K. D. (2012). Applied nucleation as a forest restoration strategy. Forest Ecology and Management, 265, 37–46.
Folke, C., Carpenter, S., Elmqvist, T., Gunderson, L., Holling, C. S., & Walker, B. (2002). Resilience and sustainable development: building adaptive capacity in a world of transformations. AMBIO: A journal of the human environment, 31(5), 437-440.
Gómez-Aparicio, L., Zamora, R., Gómez, J. M., Hódar, J. A., Castro, J., & Baraza, E. (2004). Applying plant facilitation to forest restoration: a meta-analysis of the use of shrubs as nurse plants. Ecological Applications, 14(4), 1128–1138.
Kéfi, S., Holmgren, M., & Scheffer, M. (2016). When can positive interactions cause alternative stable states in ecosystems?. Functional Ecology, 30(1), 88-97.
May, R. M. (1977). Thresholds and breakpoints in ecosystems with a multiplicity of stable states. Nature, 269(5628), 471-477.
Miriti, M. N. (2006). Ontogenetic shift from facilitation to competition in a desert shrub. Journal of Ecology, 94(5), 973–979.
Padilla, F. M., & Pugnaire, F. I. (2006). The role of nurse plants in the restoration of degraded environments. Frontiers in Ecology and the Environment, 4(4), 196–202.
Scheffer, M. 2009. Alternative Stable States. In: Critical Transitions in Nature and Society, Chapter II, pp. 11-36, Princenton University Press.
Scheffer, M., Hosper, S. H., Meijer, M. L., Moss, B., & Jeppesen, E. (1993). Alternative equilibria in shallow lakes. Trends in ecology & evolution, 8(8), 275-279.
Silliman, B. R., Schrack, E., He, Q., Cope, R., Santoni, A., Van Der Heide, T., Jacobi, M., & Van De Koppel, J. (2015). Facilitation shifts paradigms and can amplify coastal restoration efforts. Proceedings of the National Academy of Sciences, 112(46), 14295-14300.
Suding, K. N., Gross, K. L., & Houseman, G. R. (2004). Alternative states and positive feedbacks in restoration ecology. Trends in ecology & evolution, 19(1), 46-53.
A importância de diferentes escalas na dinâmica de comunidades em sistemas parasita-hospedeiro
Giovanni Cardoso dos Santos Correia
Comunidades podem ser entendidas como o conjunto de espécies ocupando o mesmo local (Hubert et al., 2015). Algo a ser considerado é que o “local” pode ser uma grande variedade de sistemas e escalas, como subamostras de uma matriz contínua (e.g. 1ha em uma floresta) a manchas de hábitat (e.g. cadáveres de baleias no mar profundo) (Hubert et al., 2015). Apesar disso, muitas das teorias existentes em ecologia de comunidades assumem que comunidades locais são sistemas fechados e isolados (Leibold et al., 2004). Porém, dinâmicas ecológicas que ocorrem em uma comunidade local não são necessariamente independentes das que ocorrem em outra comunidade local, e há processos ecológicos que podem ocorrer em escalas maiores do que a escala da comunidade local (Leibold, 2011). Portanto, para entender todos os processos ocorrendo em uma comunidade, é necessário olhar também para a metacomunidade. Metacomunidades podem ser entendidas como um conjunto de comunidades locais que são conectadas por pelo menos um de seus componentes (Leibold et al., 2004). Esse aumento na escala do estudo de comunidades nos leva a considerar como a dispersão entre comunidades locais afeta as dinâmicas ecológicas em uma comunidade local (Leibold, 2011).
A capacidade de dispersão é um processo importante quando se pensa em metacomunidades, e tanto a estruturação do hábitat quanto características do organismo possuem grande influência sobre essa capacidade (Heino et al., 2015). Por décadas foram utilizadas teorias exclusivamente sobre o local para explicar a composição de uma comunidade (Vellend, 2010). Apesar disso, mesmo pesquisadores que são considerados criadores da visão “nichocêntrica” em comunidades estavam atentos a processos que não consideravam apenas a comunidade local (Leibold & Chase, 2017). Hutchinson (1959), ao estudar insetos da família Corixidae em poças de água doce, comentou que Corixa dentipes poderia ser uma espécie “fugitiva”, por ser uma pior competidora localmente, mas possuir maior capacidade de dispersão. Outro pesquisador importante para a teoria dos nichos, MacArthur, também prestou muita atenção para a dispersão como um processo importante em comunidades. Junto com Wilson, formulou a Teoria da Biogeografia de Ilhas (MacAthur & Wilson 1963; Wilson & MacArthur, 1967), teoria em que os processos básicos para a formação de comunidades são a dispersão e a extinção. Durante os anos 70 e 80, estudos sobre processos espaciais foram deixados de lado pela maioria dos ecologistas (Leibold & Chase, 2017). Entretanto, nos anos 90 e 2000, houve um renascimento no interesse das relações entre processos espaciais e composição de comunidades, tendo a Teoria Neutra de Hubbell (2001), como seu maior estímulo (Leibold & Chase, 2017).
Mas quais são os padrões esperados quando se estuda metacomunidades? Leibold et al. (2004) reconhecem 4 principais padrões de dinâmica em metacomunidades: a dinâmica de manchas, onde existem manchas de hábitat idênticas e comunidades com interações interespecíficas determinísticas (ex. Uma espécie A que é predadora da espécie B, e A sempre leva a espécie B à extinção quando chega a uma mancha). Devido à capacidade de dispersão diferente entre a espécie A e a espécie B, é possível haver coexistência entre as duas espécies quando se analisa a metacomunidade, pois se a espécie B for mais eficiente na dispersão do que a espécie A, haverá manchas de hábitat em que a espécie A ainda não alcançou, mas já há uma população estável da espécie B. A seleção de espécies, em que as manchas de hábitat em uma região são diferentes entre si, o que permite que espécies possuam fitness diferente em cada mancha de hábitat. Apesar da heterogeneidade espacial possuir um papel importante nesse padrão de metacomunidade, a dispersão também é um processo importante, principalmente por permitir recolonização de comunidades locais após distúrbios e para conectar manchas de hábitat semelhantes, quando essas não são adjacentes. O terceiro padrão que pode ser encontrado é o efeito de massa, onde, em uma metacomunidade com manchas de hábitat diferentes, espécies consigam estar presentes em comunidades locais mesmo que as interações interespecíficas eventualmente fossem levar essa espécie à extinção. Isso ocorre se a chegada de indivíduos da espécie em uma localidade seja maior do que o número de mortes devido às interações, criando relações fonte-sumidouro entre diferentes manchas de hábitat na metacomunidade. O último padrão que os autores evidenciam é o neutro, onde as relações de nicho apresentam pouca ou nenhuma importância, e os processos mais importantes para a dinâmica da comunidade são dispersão e extinção.
Em minha pesquisa, irei trabalhar com comunidade de parasitas de peixes de riachos. Os riachos já foram previamente escolhidos, e são representantes de microbacias de terceira ordem presentes na bacia do Alto Tietê. Microbacias são bons sistemas para estudar metacomunidades, pois estão inseridas em uma matriz ambiental que é inóspita para organismos aquáticos, o que facilita a definição de comunidades locais (Heino et al., 2015), apresentam variáveis abióticas bem definidas, um pool de espécies local e regional facilmente separável e heterogeneidade ambiental considerável (Heino, 2013). Em relação aos hospedeiros, há evidência na literatura de que gradientes espaciais e ambientais são importantes para sua distribuição espacial e dinâmica das comunidades; Vieira et al. (2020) encontraram que, para ictiofauna neotropical, os padrões mais adequados para a análise de metacomunidades são o de efeito de massa e a seleção de espécies. Há a chance do padrão de metacomunidade ser o mesmo tanto para o hospedeiro quanto para o parasita, mas essa não pode ser a única explicação, devido a diferenças de escala.
Em interações simbióticas, como a relação parasita-hospedeiro, há três escalas possíveis de estudo de comunidades (Bush et al., 1997, Poulin, 2007): Infracomunidades, que são a comunidade de parasitas presentes em um determinado hospedeiro e em um determinado tempo; comunidade de componentes, que são todas as infracomunidades presentes em um subconjunto dos hospedeiros (ex. hospedeiros em uma localidade X, hospedeiros da espécie Y durante sua fase larval); e supracomunidades, que são todos os indivíduos em todas as fases de desenvolvimento em um determinado tempo e espaço. Cada uma dessas três escalas, apesar de estarem relacionadas, possuem estrutura e dinâmica diferentes, devido principalmente à diferença do tempo de existência de cada uma delas (Poulin, 2007); enquanto uma infracomunidade está restrita ao tempo de vida do indivíduo que é hospedeiro, comunidades de componentes e supracomunidades podem durar enquanto as populações ou comunidades de hospedeiros existirem. O conceito de metacomunidades permite estudar as diferentes escalas de comunidades tanto no nível do parasita quanto no nível do hospedeiro, e também permite considerar os dois níveis simultaneamente (Brown et al., 2019).
Boa parte dos estudos de comunidade de parasitas são feitos examinando padrões em um ou poucos indivíduos de uma espécie de hospedeiro (Poulin, 2007). Mas é importante considerar que parasitas podem ter ciclo de vida complexo, passando obrigatoriamente por mais de um hospedeiro durante sua vida. É possível que cada um dos hospedeiros tenha capacidades de dispersão diferentes dentro das microbacias. Como exemplo, pode ser difícil para um peixe transitar entre diferentes microbacias, seja devido à baixa capacidade dispersão, seja devido à heterogeneidade ambiental. Mas para uma ave que utiliza os riachos para alimentação, diferentes microbacias podem ser facilmente alcançáveis. Caso um parasita utilize ambos os organismos (o peixe e a ave) como hospedeiros, a capacidade de dispersão do parasita deve considerar a capacidade de dispersão de ambos os hospedeiros. Portanto, ferramentas de estudo de metacomunidades são importantes para entender a dinâmica das comunidades de parasitas, pois certas propriedades dessas comunidades podem aparecer apenas quando se considera escalas diferentes como a escala local. Além disso, é importante ter em mente como as diferentes escalas apresentam diferentes estruturas espaciais, e como essas diferentes escalas interagem para criar padrões de estrutura e dinâmica de comunidades.
Referências bibliográficas
BROWN, J. J. et al. Metacommunity theory for transmission of heritable symbionts within insect communities. Ecology and evolution, v. 10, n. 3, p. 1703-1721, 2020.
BUSH, A. O. et al. Parasitology meets ecology on its own terms: Margolis et al. revisited. The Journal of parasitology, p. 575-583, 1997.
HEINO, J. The importance of metacommunity ecology for environmental assessment research in the freshwater realm. Biological Reviews, v. 88, n. 1, p. 166-178, 2013.
HEINO, J. et al. A comparative analysis of metacommunity types in the freshwater realm. Ecology and Evolution, v. 5, n. 7, p. 1525-1537, 2015.
HUBBELL, S. P. The unified neutral theory of biodiversity and biogeography. Princeton University Press, 2001 . HUBERT, N. et al. Metacommunity speciation models and their implications for diversification theory. Ecology letters, v. 18, n. 8, p. 864-881, 2015.
HUTCHINSON, G. E. Homage to Santa Rosalia or why are there so many kinds of animals?. The American Naturalist, v. 93, n. 870, p. 145-159, 1959.
LEIBOLD, M. A. et al. The metacommunity concept: a framework for multi‐scale community ecology. Ecology letters, v. 7, n. 7, p. 601-613, 2004.
LEIBOLD, M. A. The metacommunity concept and its theoretical underpinnings. The theory of ecology, p. 163-184, 2011.
LEIBOLD, M. A.; CHASE, J. M. Metacommunity ecology, volume 59. Princeton University Press, 2017.
MACARTHUR, R. H.; WILSON, E. O. An equilibrium theory of insular zoogeography. Evolution, p. 373-387, 1963.
POULIN, R. Evolutionary ecology of parasites. Princeton University Press, 2007.
VELLEND, M. Conceptual synthesis in community ecology. The Quarterly review of biology, v. 85, n. 2, p. 183-206, 2010.
VIEIRA, T. B. et al. Elements of fish metacommunity structure in Neotropical freshwater streams. Ecology and Evolution, v. 10, n. 21, p. 12024-12035, 2020.
WILSON, E. O.; MACARTHUR, R. H. The theory of island biogeography. Princeton, NJ: Princeton University Press, 1967.
